Синтетический период митоза. митоз - фазы митоза
Временной ход митоза и цитокинеза, типичный для клетки млекопитающего. Точные цифры для разных клеток различны. Цитокинез берёт своё начало в анафазе и завершается, как правило,
к окончанию телофазы
Фаза клеточного цикла, соответствующая делению клетки, называется М-фазой. М-фазу условно подразделяют на шесть стадий, постепенно и непрерывно переходящих одна в другую. Первые пять профаза, прометафаза, метафаза, анафаза и телофаза составляют митоз, а берущий своё начало в анафазе процесс разделения цитоплазмы клетки, или цитокинез, протекает вплоть до завершения митотического цикла и, как правило, рассматривается в составе телофазы.
Длительность отдельных стадий различна и варьируется в зависимости от типа ткани, физиологического состояния организма, внешних факторов. Наиболее продолжительны стадии сопряженные с процессами внутриклеточного синтеза: профаза и телофаза. Наиболее быстротечны фазы митоза, в ходе которых происходит движение хромосом: метафаза и анафаза. Непосредственно процесс расхождения хромосом к полюсам обычно не превышает 10 минут.
Профаза

К основным событиям профазы относят конденсацию хромосом внутри ядра и образование веретена деления в цитоплазме клетки. Распад ядрышка в профазе является характерной, но не обязательной для всех клеток особенностью.
Условно за начало профазы принимается момент возникновения микроскопически видимых хромосом вследствие конденсации внутриядерного хроматина. Уплотнение хромосом происходит за счёт многоуровневой спирализации ДНК. Данные изменения сопровождаются повышением активности фосфорилаз, модифицирующих гистоны, непосредственно участвующие в компоновке ДНК. Как следствие, резко снижается транскрипционная активность хроматина, инактивируются ядрышковые гены, большая часть ядрышковых белков диссоциирует. Конденсирующиеся сестринские хроматиды в ранней профазе остаются спаренными по всей своей длине с помощью белков-когезинов, однако к началу прометафазы связь между хроматидами сохраняется лишь в области центромер. К поздней профазе на каждой центромере сестринских хроматид формируются зрелые кинетохоры необходимые хромосомам для присоединения к микротрубочкам веретена деления в прометафазе.
Наряду с процессами внутриядерной конденсации хромосом в цитоплазме начинает формироваться митотическое веретено одна из главных структур аппарата клеточного деления, ответственная за распределение хромосом между дочерними клетками. В образовании веретена деления у всех эукариотических клеток принимают участие полярные тельца, микротрубочки и кинетохоры хромосом.
С началом формирования митотического веретена в профазе сопряжены разительные изменения динамических свойств микротрубочек. Время полужизни средней микротрубочки уменьшается примерно в 20 раз от 5 минут до 15 секунд. Однако скорость их роста увеличивается примерно в 2 раза по сравнению с теми же интерфазными микротрубочками. Полимеризующиеся плюс-концы являются «динамически нестабильными» и резко переходят от равномерного роста к быстрому укорочению, при котором часто деполимеризуется вся микротрубочка. Примечательно, что для правильного функционирования митотического веретена необходим определенный баланс между процессами сборки и деполимеризации микротрубочек, так как ни стабилизированные, ни деполимеризованные микротрубочки веретена не в состоянии перемещать хромосомы.
Наряду с наблюдаемыми изменениями динамических свойств микротрубочек, слагающих нити веретена, в профазе закладываются полюса деления. Реплицированные в S-фазе центросомы расходятся в противоположных направлениях за счёт взаимодействия полюсных микротрубочек, растущих навстречу друг другу. Своими минус-концами микротрубочки погружены в аморфное вещество центросом, а процессы полимеризации протекают со стороны плюс-концов, обращенных к экваториальной плоскости клетки. При этом вероятный механизм расхождения полюсов объясняется следующим образом: динеино-подобные белки ориентируют в параллельном направлении полимеризующиеся плюс-концы полюсных микротрубочек, а кинезино-подобные белки в свою очередь расталкивают их в направлении к полюсам деления.
Параллельно конденсации хромосом и формированию митотического веретена, во время профазы происходит фрагментация эндоплазматического ретикулума, который распадается на мелкие вакуоли, расходящиеся затем к периферии клетки. Одновременно рибосомы теряют связи с мембранами ЭПР. Цистерны аппарата Гольджи также меняют свою околоядерную локализацию, распадаясь на отдельные диктиосомы, без особого порядка распределенные в цитоплазме.
Прометафаза

Прометафаза
Окончание профазы и наступление прометафазы, как правило, знаменуется распадом ядерной мембраны. Целый ряд белков ламины фосфорилируется, вследствие чего ядерная оболочка фрагментируется на мелкие вакуоли, а поровые комплексы исчезают. После разрушения ядерной мембраны хромосомы без особого порядка располагаются в области ядра. Однако вскоре все они приходят в движение.
В прометафазе наблюдается интенсивное, но беспорядочное перемещение хромосом. Поначалу отдельные хромосомы стремительно дрейфуют к ближайшему полюсу митотического веретена со скоростью, достигающей 25 мкм/мин. Вблизи полюсов деления повышается вероятность взаимодействия новосинтезированных плюс-концов микротрубочек веретена с кинетохорами хромосом. В результате такого взаимодействия кинетохорные микротрубочки стабилизируются от спонтанной деполимеризации, а их рост отчасти обеспечивает отдаление соединенной с ними хромосомы в направлении от полюса к экваториальной плоскости веретена. С другой стороны хромосому настигают тяжи микротрубочек, идущие от противоположного полюса митотического веретена. Взаимодействуя с кинетохором, они также участвуют в движении хромосомы. В результате сестринские хроматиды оказываются связанными с противоположными полюсами веретена. Усилие, развиваемое микротрубочками от разных полюсов, не только стабилизирует взаимодействие этих микротрубочек с кинетохорами, но также, в конечном счёте, приводит каждую хромосому в плоскость метафазной пластинки.
В клетках млекопитающих прометафаза протекает, как правило, в течение 10-20 минут. В нейробластах кузнечика данная стадия занимает всего 4 минуты, а в эндосперме Haemanthus и в фибробластах тритона около 30 минут.
Метафаза
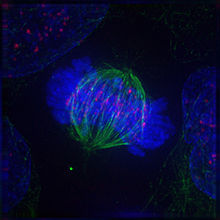
Метафаза
В завершении прометафазы хромосомы располагаются в экваториальной плоскости веретена примерно на равном расстоянии от обоих полюсов деления, образуя метафазную пластинку. Морфология метафазной пластинки в клетках животных, как правило, отличается упорядоченным расположением хромосом: центромерные участки обращены к центру веретена, а плечи к периферии клетки. В растительных клетках хромосомы зачастую лежат в экваториальной плоскости веретена без строгого порядка.
Метафаза занимает значительную часть периода митоза, и отличается относительно стабильным состоянием. Все это время хромосомы удерживаются в экваториальной плоскости веретена за счёт сбалансированных сил натяжения кинетохорных микротрубочек, совершая колебательные движения с незначительной амплитудой в плоскости метафазной пластинки.
В метафазе, также как и в течение других фаз митоза, продолжается активное обновление микротрубочек веретена путём интенсивной сборки и деполимеризации молекул тубулина. Несмотря на некоторую стабилизацию пучков кинетохорных микротрубочек, происходит постоянная переборка межполюсных микротрубочек, численность которых в метафазе достигает максимума.
К окончанию метафазы наблюдается чёткое обособление сестринских хроматид, соединение между которыми сохраняется лишь в центромерных участках. Плечи хроматид располагаются параллельно друг другу, и становится отчетливо заметной разделяющая их щель.
Анафаза

Анафаза самая короткая стадия митоза, которая начинается внезапным разделением и последующим расхождением сестринских хроматид в направлении противоположных полюсов клетки. Хроматиды расходятся с равномерной скоростью достигающей 0,5-2 мкм/мин., при этом они часто принимают V-образную форму. Их движение обусловлено воздействием значительных сил, оценочно 10 дин на хромосому, что в 10 000 раз превышает усилие, необходимое для простого продвижения хромосомы через цитоплазму с наблюдаемой скоростью.
Как правило, расхождение хромосом в анафазе состоит из двух относительно независимых процессов называемых анафазой А и анафазой В.
Анафаза А характеризуется расхождением сестринских хроматид к противоположным полюсам деления клетки. За их движение при этом отвечают те же силы, что ранее удерживали хромосомы в плоскости метафазной пластинки. Процесс расхождения хроматид сопровождается сокращением длины деполимеризующихся кинетохорных микротрубочек. Причем их распад наблюдается преимущественно в области кинетохоров, со стороны плюс-концов. Вероятно, деполимеризация микротрубочек у кинетохоров либо в области полюсов деления является необходимым условием для перемещения сестринских хроматид, так как их движение прекращается при добавлении таксола или тяжёлой воды, оказывающих стабилизирующее воздействие на микротрубочки. Механизм, лежащий в основе расхождения хромосом в анафазе А, пока остается неизвестным.
Во время анафазы В расходятся сами полюса деления клетки, и, в отличии от анафазы А, данный процесс происходит за счёт сборки полюсных микротрубочек со стороны плюс-концов. Полимеризующиеся антипараллельные нити веретена при взаимодействии отчасти и создают расталкивающее полюса усилие. Величина относительного перемещения полюсов при этом, также как и степень перекрывания полюсных микротрубочек в экваториальной зоне клетки сильно варьирует у особей разных видов. Помимо расталкивающих сил, на полюса деления воздействуют тянущие силы со стороны астральных микротрубочек, которые создаются в результате взаимодействия с динеино-подобными белками на плазматической мембране клетки.
Последовательность, продолжительность и относительный вклад каждого из двух процессов, слагающих анафазу, могут быть крайне различны. Так в клетках млекопитающих анафаза В начинается сразу вслед за началом расхождения хроматид к противоположным полюсам и продолжается вплоть до удлинения митотического веретена в 1,5-2 раза по сравнению с метафазным. В некоторых других клетках анафаза В начинается только после того как хроматиды достигают полюсов деления. У некоторых простейших в процессе анафазы В веретено удлиняется в 15 раз по сравнению с метафазным. В растительных клетках анафаза В отсутствует.
Телофаза

Телофаза
Телофаза рассматривается как заключительная стадия митоза; за её начало принимается момент остановки разделённых сестринских хроматид у противоположных полюсов деления клетки. В ранней телофазе наблюдается деконденсация хромосом и, следовательно, увеличение их в объёме. Вблизи сгруппированных индивидуальных хромосом начинается слияние мембранных пузырьков, что дает начало реконструкции ядерной оболочки. Материалом для построения мембран новообразованных дочерних ядер служат фрагменты изначально распавшейся ядерной мембраны материнской клетки, а также элементы эндоплазматического ретикулума. При этом отдельные пузырьки связываются с поверхностью хромосом и сливаются воедино. Постепенно восстанавливается наружная и внутренняя ядерные мембраны, восстанавливаются ядерная ламина и ядерные поры. В процессе восстановления ядерной оболочки дискретные мембранные пузырьки, вероятно, соединяются с поверхностью хромосом без распознавания специфических последовательностей нуклеотидов, так как в результате проведенных экспериментов было выявлено, что восстановление ядерной мембраны происходит вокруг молекул ДНК, заимствованных у любого организма, даже у бактериального вируса. Внутри заново сформировавшихся клеточных ядер хроматин переходит в дисперсное состояние, возобновляется синтез РНК, и становятся различимыми ядрышки.
Параллельно с процессами образования ядер дочерних клеток в телофазе начинается и заканчивается разборка микротрубочек веретена деления. Деполимеризация протекает в направлении от полюсов деления к экваториальной плоскости клетки, от минус-концов к плюс-концам. При этом дольше всего сохраняются микротрубочки в средней части веретена деления, которые образуют остаточное тельце Флеминга.
Окончание телофазы преимущественно совпадает с разделением тела материнской клетки цитокинезом. При этом образуются две или более дочерние клетки. Процессы, ведущие к разделению цитоплазмы, берут свое начало еще в середине анафазы и могут продолжаться после завершения телофазы. Митоз не всегда сопровождается разделением цитоплазмы, поэтому цитокинез не классифицируется в качестве отдельной фазы митотического деления и обычно рассматривается в составе телофазы.
Различают два основных типа цитокинеза: деление поперечной перетяжкой клетки и деление путём образования клеточной пластинки. Плоскость деления клетки детерминируется положением митотического веретена и проходит под прямым углом к длинной оси веретена.
При делении поперечной перетяжкой клетки место разделения цитоплазмы закладывается предварительно ещё в период анафазы, когда в плоскости метафазной пластинки под мембраной клетки возникает сократимое кольцо из актиновых и миозиновых филаментов. В дальнейшем, вследствие активности сократимого кольца, образуется борозда деления, которая постепенно углубляется вплоть до полного разделения клетки. По окончании цитокинеза сократимое кольцо полностью распадается, а плазматическая мембрана стягивается вокруг остаточного тельца Флеминга, состоящего из скопления остатков двух групп полюсных микротрубочек, тесно упакованных вместе с материалом плотного матрикса.
Деление путём образования клеточной пластинки начинается с перемещения мелких ограниченных мембраной пузырьков по направлению к экваториальной плоскости клетки. Здесь они сливаются, образуя дисковидную, окружённую мембраной структуру раннюю клеточную пластинку. Мелкие пузырьки происходят в основном из аппарата Гольджи и перемещаются к экваториальной плоскости вдоль остаточных полюсных микротрубочек веретена деления, образующих цилиндрическую структуру, называемую фрагмопластом. По мере расширения клеточной пластинки микротрубочки раннего фрагмопласта попутно перемещаются к периферии клетки, где за счёт новых мембранных пузырьков продолжается рост клеточной пластинки вплоть до её окончательного слияния с мембраной материнской клетки. После окончательного разделения дочерних клеток в клеточной пластинке откладываются микрофибриллы целлюлозы, завершая образование жёсткой клеточной стенки.
| Прево, Жан-Луи |
Наследственность как всеобщее свойство живых организмов тесно связана с другим важнейшим свойством живого - размножением . Благодаря размножению осуществляется преемственность между родительскими особями и их потомством. В основе размножения лежит процесс деления клеток.
Хромосомы: индивидуальность, парность, число
Во время деления клетки хорошо заметны хромосомы. При изучении хромосом разных видов живых организмов было обнаружено, что их набор строго индивидуален. Это касается числа, формы, черт строения и величины хромосом. Набор хромосом в клетках тела, характерный для данного вида растений, животных, называется кариотипом.
В любом многоклеточном организме существует два вида клеток - соматические (клетки тела) и половые клетки, или гаметы. В половых клетках число хромосом в 2 раза меньше, чем в соматических. В соматических клетках все хромосомы представлены парами - такой набор называется диплоидным и обозначается 2/1- Парные хромосомы (одинаковые по величине, форме, строению) называются гомологичными.
В половых клетках каждая из хромосом находится в одинарном числе. Такой набор называется гаплоидным и обозначается n.
Митоз. Подготовка клетки к делению
Наиболее распространенным способом деления соматических клеток является митоз. Во время митоза клетка проходит ряд последовательных стадий, или фаз, в результате которых каждая дочерняя клетка получает такой же набор хромосом, какой был у материнской клетки.
Во время подготовки клетки к делению - в период интерфазы (период между двумя актами деления) число хромосом удваивается. Вдоль каждой исходной хромосомы из имеющихся в клетке химических соединений синтезируется ее точная копия. Удвоенная хромосома состоит из двух половинок - хроматид. Каждая из хроматид содержит одну молекулу ДНК- В период интерфазы в клетке происходит процесс биосинтеза белка, удваиваются также все важнейшие структуры клетки. Продолжительность интерфазы в среднем 10-20 ч. Затем наступает процесс деления клетки - митоз.
Фазы митоза
Во время митоза клетка проходит следующие четыре фазы: профаза, метафаза, анафаза, телофаза.
В профазе хорошо видны ценгриоли - органоиды, играющие определенную роль в делении дочерних хромосом. Центриолй делятся и расходятся к разным полюсам. От них протягиваются нити, образующие веретено деления, которое регулирует расхождение хромосом к полюсам делящейся клетки. В конце профазы ядерная оболочка распадается, исчезает ядрышко, хромосомы спирализуются и укорачиваются.
Метафаза характеризуется наличием хорошо видимых хромосом, располагающихся в экваториальной плоскости клетки. Каждая хромосома состоит из двух хроматид и имеет перетяжку - центромеру, к которой прикрепляются нити веретена деления. После деления центромеры каждая хроматида становится самостоятельной дочерней хромосомой.
В анафазе дочерние хромосомы расходятся к разным полюсам клетки.
В последней стадии - телофазе - хромосомы вновь раскручиваются и приобретают вид длинных тонких нитей. Вокруг них возникает ядерная оболочка, в ядре формируется ядрышко.
В процессе деления цитоплазмы все ее органоиды равномерно распределяются между дочерними клетками. Весь процесс митоза продолжается обычно 1-2 ч.
В результате митоза все дочерние клетки содержат одинаковый набор хромосом и одни и те же гены. Следовательно, митоз - это способ деления клетки, заключающийся в точном распределении генетического материала между дочерними клетками, обе дочерние клетки получают диплоидный набор хромосом.
Лекция № 10
Количество часов: 2
МИТОЗ
1. Жизненный цикл клетки
2. Митоз. Стадии митоза, их продолжительность и характеристика
3. Амитоз. Эндорепродукция
1. Жизненный цикл клетки
Клетки многоклеточного организма чрезвычайно разнообразны по выполняемым функциям. В соответствии со специализацией клетки имеют разную продолжительность жизни. Так нервные клетки после завершения эмбриогенеза перестают делиться и функционируют на протяжении всей жизни организма. Клетки же других тканей (костного мозга, эпидермиса, эпителия тонкого кишечника) в процессе выполнения своей функции быстро погибают и замещаются новыми в результате клеточного деления. Деление клеток лежит в основе развития, роста и размножения организмов. Деление клеток также обеспечивает самообновление тканей на протяжении жизни организма и восстановление их целостности после повреждения. Существует два способа деления соматических клеток: амитоз и митоз . Преимущественно распространено непрямое деление клеток (митоз). Размножение с помощью митоза называют бесполым размножением, вегетативным размножением или клонированием.
Жизненный цикл клетки (клеточный цикл) – это существование клетки от деления до следующего деления или смерти. Продолжительность клеточного цикла в размножающихся клетках составляет 10-50 ч и зависит от типа клеток, их возраста, гормонального баланса организма, температуры и других факторов. Детали клеточного цикла варьируют среди разных организмов. У одноклеточных организмов жизненный цикл совпадает с жизнью особи. В непрерывно размножающихся тканевых клетках клеточный цикл совпадает с митотическим циклом.
Митотический цикл - совокупность последовательных и взаимосвязанных процессов в период подготовки клетки к делению и период деления (рис 1). В соответствие с приведенным выше определением митотический цикл подразделяют на интерфазу и митоз (греч. “митос” - нить).
Интерфаза - период между двумя делениями клетки - подразделяется на фазы G 1 , S и G 2 (ниже указана их продолжительность, типичная для растительных и животных клеток.). По продолжительности интерфаза составляет большую часть митотического цикла клетки. Наиболее вариабельны по времени G 1 и G 2 -периоды.
G 1 (от англ. grow – расти, увеличиваться). Продолжительность фазы составляет 4–8 ч. Это фаза начинается сразу после образования клетки. В этой фазе в клетке усиленно синтезируются РНК и белки, повышается активность ферментов, участвующих в синтезе ДНК. Если клетка в дальнейшем не делится, то переходит в фазу G 0 – период покоя. С учетом периода покоя клеточный цикл может длиться недели или даже месяцы (клетки печени).
S (от англ. synthesis - синтез). Длительность фазы составляет 6–9 ч. Масса клетки продолжает увеличиваться, и происходит удвоение хромосомной ДНК. Две спирали старой молекулы ДНК расходятся, и каждая становится матрицей для синтеза новых цепей ДНК. В результате каждая из двух дочерних молекул обязательно включает одну старую спираль и одну новую. Тем не менее хромосомы остаются одинарными по структуре, хотя и удвоенными по массе, так как две копии каждой хромосомы (хроматиды) все еще соединены друг с другом по всей длине. После завершения фазы S митотического цикла клетка не сразу начинает делиться.
G 2 . В этой фазе в клетке завершается процесс подготовки к митозу: накапливается АТФ, синтезируются белки ахроматинового веретена, удваиваются центриоли. Масса клетки продолжает увеличиваться до тех пор, пока она приблизительно вдвое не превысит начальную, а затем наступает митоз.
Рис. Митотический цикл: М - митоз, П - профаза, Мф - метафаза, А - анафаза, Т- телофаза, G 1 - пресинтетический период, S - синтетический период, G 2 - постсинтетический
2. Митоз. Стадии митоза, их продолжительность и характеристика. Митоз условно разделяют на четыре фазы: профазу, метафазу, анафазу и телофазу.
Профаза. Две центриоли начинают расходиться к противоположным полюсам ядра. Ядерная мембрана разрушается; одновременно специальные белки объединяются, формируя микротрубочки в виде нитей. Центриоли, расположенные теперь на противоположных полюсах клетки, оказывают организующее воздействие на микротрубочки, которые в результате выстраиваются радиально, образуя структуру, напоминающую по внешнему виду цветок астры («звезда»). Другие нити из микротрубочек протягиваются от одной центриоли к другой, образуя веретено деления. В это время хромосомы спирализуются и вследствие этого утолщаются. Они хорошо видны в световом микроскопе, особенно после окрашивания. Считывание генетической информации с молекул ДНК становится невозможным: синтез РНК прекращается, ядрышко исчезает. В профазе хромосомы расщепляются, но хроматиды все еще остаются скрепленными попарно в зоне центромеры. Центромеры тоже оказывают организующее воздействие на нити веретена, которые теперь тянутся от центриоли к центромере и от нее к другой центриоли.
Метафаза. В метафазе спирализация хромосом достигает максимума, и укороченные хромосомы устремляются к экватору клетки, располагаясь на равном расстоянии от полюсов. Образуется экваториальная, или метафазная, пластинка. На этой стадии митоза отчетливо видна структура хромосом, их легко сосчитать и изучить их индивидуальные особенности. В каждой хромосоме имеется область первичной перетяжки - центромера, к которой во время митоза присоединяются нить веретена деления и плечи. На стадии метафазы хромосома состоит из двух хроматид, соединенных между собой только в области центромеры.

Рис. 1. Митоз растительной клетки. А -
интерфаза;
Б, В, Г, Д-
профаза; Е,
Ж-метафаза; 3, И - анафаза; К, Л,
М-телофаза
В анафазе вязкость цитоплазмы уменьшается, центромеры разъединяются, и с этого момента хроматиды становятся самостоятельными хромосомами. Нити веретена деления, прикрепленные к центромерам, тянут хромосомы к полюсам клетки, а плечи хромосом при этом пассивно следуют за центромерой. Таким образом, в анафазе хроматиды удвоенных еще в интерфазе хромосом точно расходятся к полюсам клетки. В этот момент в клетке находятся два диплоидных набора хромосом (4n4с).
Таблица 1. Митотический цикл и митоз
| Фазы | Процесс, происходящий в клетке |
|
| Интерфаза | Пресинтетический период (G1) | Синтез белка. На деспирализованных молекулах ДНК синтезируется РНК |
| Синтетический период (S) | Синтез ДНК - самоудвоение молекулы ДНК. Построение второй хроматиды, в которую переходит вновь образовавшаяся молекула ДНК: получаются двухроматидные хромосомы |
|
| Постсинтетический период (G2) | Синтез белка, накопление энергии, подготовка к делению |
|
| Фазы митоза | Профаза | Двухроматидные хромосомы спирализуются, ядрышки растворяются, центриоли расходятся, ядерная оболочка растворяется, образуются нити веретена деления |
| Метафаза | Нити веретена деления присоединяются к центромерам хромосом, двухроматидные хромосомы сосредоточиваются на экваторе клетки |
|
| Анафаза | Центромеры делятся, однохроматидные хромосомы растягиваются нитями веретена деления к полюсам клетки |
|
| Телофаза | Однохроматидные хромосомы деспирализуются, сформировывается ядрышко, восстанавливается ядерная оболочка, на экваторе начинает закладываться перегородка между клетками, растворяются нити веретена деления |
|
В телофазе хромосомы раскручиваются, деспирализуются. Из мембранных структур цитоплазмы образуется ядерная оболочка. В это время восстанавливается ядрышко. На этом завершается деление ядра (кариокинез), затем происходит деление тела клетки (или цитокинез). При делении животных клеток на их поверхности в плоскости экватора появляется борозда, постепенно углубляющаяся и разделяющая клетку на две половины - дочерние клетки, в каждой их которых имеется по ядру. У растений деление происходит путем образования так называемой клеточной пластинки, разделяющей цитоплазму: она возникает в экваториальной области веретена, а затем растет во все стороны, достигая клеточной стенки (т.е. растет изнутри кнаружи). Клеточная пластинка формируется из материала, поставляемого эндоплазматической сетью. Затем каждая из дочерних клеток образует на своей стороне клеточную мембрану и, наконец, на обеих сторонах пластинки образуются целлюлозные клеточные стенки. Особенности протекания митоза у животных и растений приведены в таблице 2.
Таблица 2. Особенности митоза у растений и у животных
| Растительная клетка | Животная клетка |
| Центриолей нет Звезды не образуются Образуется клеточная пластинка При цитокенезе борозда не образуется Митозы преимущественно происходят в меристемах | Центриоли имеются Звезды образуются Клеточная пластинка не образуется При цитокинезе образуется борозда Митозы происходят в различных тканях организма |
Так из одной клетки формируются две дочерние, в которых наследственная информация точно копирует информацию, содержавшуюся в материнской клетке. Начиная с первого митотического деления оплодотворенной яйцеклетки (зиготы) все дочерние клетки, образовавшиеся в результате митоза, содержат одинаковый набор хромосом и одни и те же гены. Следовательно, митоз - это способ деления клеток, заключающийся в точном распределении генетического материала между дочерними клетками. В результате митоза обе дочерние клетки получают диплоидный набор хромосом.
Весь процесс митоза занимает в большинстве случаев от 1 до 2 часов. Частота митоза в разных тканях и у разных видов различна. Например, в красном костном мозге человека, где каждую секунду образуется 10 млн эритроцитов, в каждую секунду должно происходить 10 млн. митозов. А в нервной ткани митозы крайне редки: так, в центральной нервной системе клетки в основном перестают делиться уже в первые месяцы после рождения; а в красном костном мозге, в эпителиальной выстилке пищеварительного тракта и в эпителии почечных канальцев они делятся до конца жизни.
Регуляция митоза, вопрос о пусковом механизме митоза.
Факторы, побуждающие клетку к митозу точно не известны. Но полагают, что большую роль играет фактор соотношения объемов ядра и цитоплазмы (ядерно-плазменное соотношение). По некоторым данным, отмирающие клетки продуцируют вещества, способные стимулировать деление клетки. Белковые факторы, отвечающие за переход в фазу М, первоначально были идентифицированы на основе экспериментов по слиянию клеток. Слияние клетки, находящейся в любой стадии клеточного цикла, с клеткой находящейся в М фазе, приводит к вхождению ядра первой клетки в М фазу. Это означает, что в клетке находящейся в М фазе существует цитоплазматический фактор способный активировать М фазу. Позднее этот фактор был вторично обнаружен в экспериментах по переносу цитоплазмы между ооцитами лягушки, находящимися на различных стадиях развития, и был назван "фактором созревания" MPF (maturation promoting factor). Дальнейшее изучение MPF показало, что этот белковый комплекс детерминирует все события М-фазы. На рисунке показано, что распад ядерной мембраны, конденсация хромосом, сборка веретена, цитокинез регулируются MPF.
Митоз тормозится высокой температурой, высокими дозами ионизирующей радиации, действием растительных ядов. Один из таких ядов называется колхицин. С его помощью можно остановить митоз на стадии метафазной пластинки, что позволяет подсчитать число хромосом и дать каждой из них индивидуальную характеристику, т. е. провести кариотипирование.
4. Амитоз. Эндорепродукция
Амитоз (от греч. а – отриц. частица и митоз) -прямоеделение интерфазного ядра путем перешнуровывания без преобразования хромосом. При амитозе не происходит равномерное расхождение хроматид к полюсам. И это деление не обеспечивает образование генетически равноценных ядер и клеток. По сравнению с митозом амитоз более кратковременный и экономичный процесс. Амитотическое деление может осуществляться несколькими способами. Наиболее распространенный тип амитоза – это перешнуровывание ядра на две части. Этот процесс начинается с разделения ядрышка. Перетяжка углубляется, и ядро разделяется надвое. После этого начинается разделение цитоплазмы, однако это происходит не всегда. Если амитоз ограничивается только делением ядра, то это приводит к образованию дву- и многоядерных клеток. При амитозе может также происходить почкование и фрагментация ядер.
Клетка, претерпевшая амитоз, в последующем не способна вступить в нормальный митотический цикл.
Амитоз встречается в клетках различных тканей растений и животных. У растений амитотическое деление довольно часто встречается в эндосперме, в специализирующихся клетках корешков и в клетках запасающих тканей. Амитоз также наблюдается в высокоспециализированных клетках с ослабленной жизнеспособностью или дегенерирующих, при различных патологических процессах, таких как злокачественный рост, воспаление и т. п.
Основным процессом в подготовке клетки к митозу является репликация ДНК и удвоение хромосом. Но синтез ДНК и митоз непосредственно не связаны, т.к. окончательный синтез ДНК не является непосредственной причиной вступления клетки в митоз. Поэтому в ряде случаев клетки после удвоения хромосом не делятся, ядро и все клетки увеличиваются в объеме, становятся полиплоидными. Такое явление - редупликация хромосом, без деления, выработалась в процессе эволюции как способ, обеспечивающий рост органов без увеличения числа клеток. Все случаи, когда происходит редупликация хромосом или репликация ДНК, но не наступает митоз, называются эндорепродукциями. Клетки становятся полиплоидными. Как постоянный процесс эндорепродукция наблюдается в клетках печени, эпителия мочевыводящих путей млекопитающих. В случае эндомитоза хромосомы после редупликации становятся видны, но ядерная оболочка не разрушается.
Если делящиеся клетки на некоторое время охладить или об работать их каким-либо веществом, разрушающим микротрубочки веретена (например, колхицином), то деление клеток прекратит ся. При этом исчезнет веретено, а хромосомы без расхождения к полюсам будут продолжать цикл своих превращений: они начнут набухать, одеваться ядерной оболочкой. Так возникают за счет объединения всех неразошедшихся наборов хромосом крупные новые ядра. Они, естественно, будут содержать вначале 4п число хроматид и соответственно 4с количество ДНК. По определению, это уже не диплоидная, а тетраплоидная клетка. Такие полипло идные клетки могут из стадии gi переходить в S -период и, если убрать колхицин, снова делиться митотическим путем, давая уже потомков с 4 п числом хромосом. В результате можно получить полиплоидные клеточные линии разной величины плоидности. Этот прием часто используется для получения полиплоидных растений.
Как оказалось, во многих органах и тканях нормальных ди плоидных организмов животных и растений встречаются клетки с крупными ядрами, количество ДНК в которых кратно больше 2 п. При делении таких клеток видно, что количество хромосом у них также кратно увеличено по сравнению с обычными дипло идными клетками. Эти клетки являются результатом соматиче ской полиплоидии. Часто это явление называют эндорепродук цией - - появление клеток с увеличенным содержанием ДНК. Появление подобных клеток происходит в результате отсутствия в целом или незавершенности отдельных этапов митоза. Суще ствует несколько точек в процессе митоза, блокада которых приведет к его остановке и к появлению полиплоидных клеток. Блок может наступить при переходе от С 2 -периода к собствен но митозу, остановка может произойти в профазе и метафазе, в последнем случае часто происходит нарушение целостности ве ретена деления. Наконец, нарушения цитотомии также могут пре кратить деление, что приведет к появлению двуядерных и поли плоидных клеток.
При естественной блокаде митоза в самом его начале, при переходе G 2 - профазы, клетки приступают к следующему циклу репликации, который приведет к прогрессивному увеличению ко личества ДНК в ядре. При этом не наблюдается никаких морфо логических особенностей таких ядер, кроме их больших размеров. При увеличении ядер в них не выявляются хромосомы митоти ческого типа. Часто такой тип эндорепродукции без митотической конден сации хромосом встречается у беспозвоночных животных, обнаруживается он также и у позвоночных животных, и у растений. У беспозвоночных в результате блока митоза степень поли плоидии может достигать огромных значений. Так, в гигантских нейронах моллюска тритонии, ядра которых достигают величины до 1 мм (!), содержится более 2-10 5 гаплоидных наборов ДНК. Другим примером гигантской полиплоидной клетки, образо вавшейся в результате редупликации ДНК без вступления кле ток в митоз, может служить клетка шелкоотделительной железы тутового шелкопряда. Ее ядро имеет причудливую ветвистую форму и может содержать огромные количества ДНК. Гигантские клетки железы пищевода аскариды могут содержать до 100000с ДНК.
Особый случай эндорепродукции представляет собой увеличе ние плоидности путем политении. При политении в S -периоде при репликации ДИК новые до черние хромосомы продолжают оставаться в деспирализованном состоянии, но располагаются друг около друга, не расходятся и не претерпевают митотическую конденсацию. В таком истинно интерфазном виде хромосомы снова вступают в следующий цикл репликации, снова удваиваются и не расходятся. По степенно в результате репликации и нерасхождения хромосомных нитей образуется многонитчатая, политенная структура хромосо мы интерфазного ядра. Последнее обстоятельство необходимо под черкнуть, так как такие гигантские политенные хромосомы ни когда не участвуют в митозе, более того - это истинно интерфаз ные хромосомы, участвующие в синтезе ДНК и РНК. От митотических хромосом они резко отличаются и по разме рам: в несколько раз толще митотических хромосом из-за того, что состоят из пучка множественных неразошедшихся хро матид - по объему политенные хромосомы дрозофилы в 1000 раз «больше митотических. Они в 70-250 раз длиннее митотических из-за того, что в интерфазном состоянии хромосомы менее конденсированы (спирализованы), чем митотические хромосомы. Кроме того, у двукрылых их общее число в клетках равно гаплоидному из-за того, что при политенизации происходит объединение, конъюгация гомологичных хромосом. Так, у дрозофилы в диплоидной соматической клетке 8 хромосом, а в гигантской клетке слюнной железы - 4. Встречаются гигантские полиплоидные ядра с политенными хромосомами у некоторых личинок двукрылых насекомых в клет ках слюнных желез, кишечника, мальпигиевых сосудов, жирового тела и т. д. Описаны политенные хромосомы в макронуклеусе инфузо рии стилонихии. Лучше всего этот тип эндорепродукции изучен у насекомых. Было подсчитано, что у дрозофилы в клетках слюнных желез может произойти до 6-8 циклов редупликации, что приведет к общей плоидности клетки, равной 1024. У некоторых хирономид (их личинку называют мотылем) плоидность в этих клетках до стигает 8000-32000. В клетках политенные хромосомы начинают быть видны после достижения политении в 64-128 п, до этого такие ядра ничем, кроме размера, не отличаются от окружающих диплоидных ядер.
Отличаются политенные хромосомы и своим строением: они структурно неоднородны по длине, состоят из дисков, междис ковых участков и пуфов. Рисунок расположения дисков строго характерен для каждой хромосомы и отличается даже у близких видов животных. Диски представляют собой участки конденсированного хро матина. Диски могут отличаться друг от друга по толщине. Общее их число у политенных хромосом хирономид достигает 1,5-2,5 тыс. У дрозофилы имеется около 5 тыс. дисков. Диски разделены междисковыми пространствами, состоящими, так же как и диски, из фибрилл хроматина, только более рыхла упакованных. На политенных хромосомах двукрылых часто видны вздутия, пуфы. Оказалось, что пуфы возникают на местах некоторых дис ков за счет их деконденсации и разрыхления. В пуфах выявля ется РНК, которая там же и синтезируется. Рисунок расположения и чередования дисков на политенных хромосомах постоянен и не зависит ни от органа, ни от возраста животного. Это является хорошей иллюстрацией одинаковости качества генетической информации в каждой клетке организма. Пуфы являются временными образованиями на хромосомах, и в процессе развития организма существует определенная последовательность в их появлении и исчезновении на гене тически различных участках хромосомы. Эта последо вательность различна для разных тканей. Сейчас доказано, что образование пуфов на политенных хромосомах - - это выражение генной активности: в пуфах синтезируются РНК, необходимые для проведения белковых синтезов на разных этапах развития насекомого. В естественных условиях у двукрылых особенно активны в отношении синтеза РНК два самых крупных пуфа, так называе мые кольца Бальбиани, который описал их 100 лет тому назад.
В других случаях эндорепродукции полиплоидные клетки воз никают в результате нарушений аппарата деления - веретена: при этом происходит митотическая конденсация хромосом. Такое явление носит название эндомитоз, потому что конденсация хро мосом и их изменения происходят внутри ядра, без исчезновения ядерной оболочки. Впервые явление эндомитоза было хорошо изучено в клетках: различных тканей водяного клопа - - геррии. В начале эндоми тоза хромосомы конденсируются, благодаря чему становятся хо рошо различимы внутри ядра, затем хроматиды обособляются, вытягиваются. Эти стадии по состоянию хромосом могут соответствовать профазе и метафазе обычного митоза. Затем хромосомы в таких ядрах исчезают, и ядро принимает вид обычного интер фазного ядра, но размер его увеличивается в соответствии с уве личением плоидности. После очередной редупликации ДНК такой цикл эндомитоза повторяется. В результате могут возникнуть полиплоидные (32 п) и даже гигантские ядра. Сходный тип эндомитоза описан при развитии макронуклеу сов у некоторых инфузорий, у целого ряда растений.
Результат эндорепродукции: полиплоидия и увеличение размеров клетки.
Значение эндорепродукции: не прерывается деятельность клетки. Так, например, деле ние нервных клеток привело бы к временному выключению их функций; эндорепродукция позволяет без перерыва в функциони ровании нарастить клеточную массу и тем самым увеличить объ ем работы, выполняемый одной клеткой.
увеличение продуктивности клеток.
Профаза
. В профазе происходит конденсация хромосом, и они становятся видимыми при световой микроскопии. Хромосомы по мере компактизации ДНП приобретают строение хорошо окрашивающихся нитей. Число хромосом равно 4n, что соответствует количеству ДНК 4с. В связи с инактивацией генов в области ядрышкового организатора и угнетением синтеза РНК в профазе отмечается исчезновение ядрышек. Ядерная оболочка постепенно распадается на фрагменты и мелкие мембранные пузырьки. При этом к противоположным полюсам клетки расходятся центриоли.
В сателлитном участке материнской центриоли
начинается образование микротрубочек, из которых формируются нити веретена деления.
Метафаза
. Характерным событием для метафазы является перемещение хромосом в экваториальную плоскость веретена. Здесь они располагаются строго закономерно, образуя метафазную пластинку (при взгляде на веретено деления сбоку). Если рассматривать группу метафазных хромосом со стороны полюсов веретена, то отчетливо выступает фигура, напоминающая звезду (так называемая материнская звезда). В этот период можно определить число, форму и размеры хромосом (d-хромосом, двойных хромосом), составляющих метафазную пластинку.
К концу метафазы
продольные половинки хромосом (сестринские хроматиды) обособляются на всем протяжении, кроме зоны первичной перетяжки.
Для каждого вида животных характерно строго постоянное число хромосом в соматических клетках. Для человека оно равно 46. По длине хромосом различают чередование окрашенных и неокрашенных участков. При этом каждая хромосома отличается неповторимым рисунком дифференциальной окраски. Хромосомы человека подразделяются на 7 групп по их размерам и особенностям строения (А, В, С, D, Е, F, G) и каждая хромосома имеет свой номер. Совокупность признаков строения хромосом, их размеров и числа составляет то, что называют кариотипом.
Анафаза включает процесс расхождения хромосом к полюсам делящейся клетки. Механизм движения хромосом объясняется гипотезой скользящих нитей, согласно которой состоящие из микротрубочек нити веретена, взаимодействуя друг с другом и с сократительными белками, тянут хромосомы к полюсам. Скорость движения хромосом достигает 0,2-0,5 мкм/мин, а вся анафаза продолжается 2-3 мин. Анафаза заканчивается перемещением двух идентичных наборов хромосом (s-хромосом, или одиночных хромосом) к полюсам, где они сближаются, образуя фигуры, напоминающие по внешнему виду (если смотреть со стороны полюса) звезды. Эти фигуры называют дочерними звездами.
Так как хромосомные звезды образуются у каждого из полюсов, данную стадию митоза иногда называют стадией двойной звезды (диастер), или стадией дочерних звезд. Телофаза - конечная стадия митоза, в течение которой на полюсах веретена реконструируются дочерние ядра. Перестройка телофазных хромосом напоминает процессы их изменения в профазе, но происходящие в обратном направлении. При взаимодействии хромосом с мембранными пузырьками цитоплазмы формируется ядерная оболочка. С переходом хромосом в интерфазное состояние образуются новые ядрышки. Телофаза завершается разделением тела клетки - цитотомией, или цитокинезом, что приводит к образованию двух дочерних клеток.
Часть клеток может выходить из цикла репродукции и вступить на путь дифференцировки. Некоторые клетки могут выходить из клеточного цикла в G1-периоде или после S-периода и находиться в покое (Go-период). Такие покоящиеся клетки сохраняют способность к делению и могут снова входить в цикл размножения.
Учебное видео: митоз клетки и его стадии
При проблемах с просмотром скачайте видео со страницы Оглавление темы "Строение клетки. Клеточные элементы.":Лекция № 10
Количество часов: 2
МИТОЗ
1. Жизненный цикл клетки
2. Митоз. Стадии митоза, их продолжительность и характеристика
3. Амитоз. Эндорепродукция
1. Жизненный цикл клетки
Клетки многоклеточного организма чрезвычайно разнообразны по выполняемым функциям. В соответствии со специализацией клетки имеют разную продолжительность жизни. Так нервные клетки после завершения эмбриогенеза перестают делиться и функционируют на протяжении всей жизни организма. Клетки же других тканей (костного мозга, эпидермиса, эпителия тонкого кишечника) в процессе выполнения своей функции быстро погибают и замещаются новыми в результате клеточного деления. Деление клеток лежит в основе развития, роста и размножения организмов. Деление клеток также обеспечивает самообновление тканей на протяжении жизни организма и восстановление их целостности после повреждения. Существует два способа деления соматических клеток: амитоз и митоз . Преимущественно распространено непрямое деление клеток (митоз). Размножение с помощью митоза называют бесполым размножением, вегетативным размножением или клонированием.
Жизненный цикл клетки (клеточный цикл) – это существование клетки от деления до следующего деления или смерти. Продолжительность клеточного цикла в размножающихся клетках составляет 10-50 ч и зависит от типа клеток, их возраста, гормонального баланса организма, температуры и других факторов. Детали клеточного цикла варьируют среди разных организмов. У одноклеточных организмов жизненный цикл совпадает с жизнью особи. В непрерывно размножающихся тканевых клетках клеточный цикл совпадает с митотическим циклом.
Митотический цикл - совокупность последовательных и взаимосвязанных процессов в период подготовки клетки к делению и период деления (рис 1). В соответствие с приведенным выше определением митотический цикл подразделяют на интерфазу и митоз (греч. “митос” - нить).
Интерфаза - период между двумя делениями клетки - подразделяется на фазы G 1 , S и G 2 (ниже указана их продолжительность, типичная для растительных и животных клеток.). По продолжительности интерфаза составляет большую часть митотического цикла клетки. Наиболее вариабельны по времени G 1 и G 2 -периоды.
G 1 (от англ. grow – расти, увеличиваться). Продолжительность фазы составляет 4–8 ч. Это фаза начинается сразу после образования клетки. В этой фазе в клетке усиленно синтезируются РНК и белки, повышается активность ферментов, участвующих в синтезе ДНК. Если клетка в дальнейшем не делится, то переходит в фазу G 0 – период покоя. С учетом периода покоя клеточный цикл может длиться недели или даже месяцы (клетки печени).
S (от англ. synthesis - синтез). Длительность фазы составляет 6–9 ч. Масса клетки продолжает увеличиваться, и происходит удвоение хромосомной ДНК. Две спирали старой молекулы ДНК расходятся, и каждая становится матрицей для синтеза новых цепей ДНК. В результате каждая из двух дочерних молекул обязательно включает одну старую спираль и одну новую. Тем не менее хромосомы остаются одинарными по структуре, хотя и удвоенными по массе, так как две копии каждой хромосомы (хроматиды) все еще соединены друг с другом по всей длине. После завершения фазы S митотического цикла клетка не сразу начинает делиться.
G 2 . В этой фазе в клетке завершается процесс подготовки к митозу: накапливается АТФ, синтезируются белки ахроматинового веретена, удваиваются центриоли. Масса клетки продолжает увеличиваться до тех пор, пока она приблизительно вдвое не превысит начальную, а затем наступает митоз.
Рис. Митотический цикл: М - митоз, П - профаза, Мф - метафаза, А - анафаза, Т- телофаза, G 1 - пресинтетический период, S - синтетический период, G 2 - постсинтетический
2. Митоз. Стадии митоза, их продолжительность и характеристика. Митоз условно разделяют на четыре фазы: профазу, метафазу, анафазу и телофазу.
Профаза. Две центриоли начинают расходиться к противоположным полюсам ядра. Ядерная мембрана разрушается; одновременно специальные белки объединяются, формируя микротрубочки в виде нитей. Центриоли, расположенные теперь на противоположных полюсах клетки, оказывают организующее воздействие на микротрубочки, которые в результате выстраиваются радиально, образуя структуру, напоминающую по внешнему виду цветок астры («звезда»). Другие нити из микротрубочек протягиваются от одной центриоли к другой, образуя веретено деления. В это время хромосомы спирализуются и вследствие этого утолщаются. Они хорошо видны в световом микроскопе, особенно после окрашивания. Считывание генетической информации с молекул ДНК становится невозможным: синтез РНК прекращается, ядрышко исчезает. В профазе хромосомы расщепляются, но хроматиды все еще остаются скрепленными попарно в зоне центромеры. Центромеры тоже оказывают организующее воздействие на нити веретена, которые теперь тянутся от центриоли к центромере и от нее к другой центриоли.
Метафаза. В метафазе спирализация хромосом достигает максимума, и укороченные хромосомы устремляются к экватору клетки, располагаясь на равном расстоянии от полюсов. Образуется экваториальная, или метафазная, пластинка. На этой стадии митоза отчетливо видна структура хромосом, их легко сосчитать и изучить их индивидуальные особенности. В каждой хромосоме имеется область первичной перетяжки - центромера, к которой во время митоза присоединяются нить веретена деления и плечи. На стадии метафазы хромосома состоит из двух хроматид, соединенных между собой только в области центромеры.

Рис. 1. Митоз растительной клетки. А -
интерфаза;
Б, В, Г, Д-
профаза; Е,
Ж-метафаза; 3, И - анафаза; К, Л,
М-телофаза
В анафазе вязкость цитоплазмы уменьшается, центромеры разъединяются, и с этого момента хроматиды становятся самостоятельными хромосомами. Нити веретена деления, прикрепленные к центромерам, тянут хромосомы к полюсам клетки, а плечи хромосом при этом пассивно следуют за центромерой. Таким образом, в анафазе хроматиды удвоенных еще в интерфазе хромосом точно расходятся к полюсам клетки. В этот момент в клетке находятся два диплоидных набора хромосом (4n4с).
Таблица 1. Митотический цикл и митоз
| Фазы | Процесс, происходящий в клетке |
|
| Интерфаза | Пресинтетический период (G1) | Синтез белка. На деспирализованных молекулах ДНК синтезируется РНК |
| Синтетический период (S) | Синтез ДНК - самоудвоение молекулы ДНК. Построение второй хроматиды, в которую переходит вновь образовавшаяся молекула ДНК: получаются двухроматидные хромосомы |
|
| Постсинтетический период (G2) | Синтез белка, накопление энергии, подготовка к делению |
|
| Фазы митоза | Профаза | Двухроматидные хромосомы спирализуются, ядрышки растворяются, центриоли расходятся, ядерная оболочка растворяется, образуются нити веретена деления |
| Метафаза | Нити веретена деления присоединяются к центромерам хромосом, двухроматидные хромосомы сосредоточиваются на экваторе клетки |
|
| Анафаза | Центромеры делятся, однохроматидные хромосомы растягиваются нитями веретена деления к полюсам клетки |
|
| Телофаза | Однохроматидные хромосомы деспирализуются, сформировывается ядрышко, восстанавливается ядерная оболочка, на экваторе начинает закладываться перегородка между клетками, растворяются нити веретена деления |
|
В телофазе хромосомы раскручиваются, деспирализуются. Из мембранных структур цитоплазмы образуется ядерная оболочка. В это время восстанавливается ядрышко. На этом завершается деление ядра (кариокинез), затем происходит деление тела клетки (или цитокинез). При делении животных клеток на их поверхности в плоскости экватора появляется борозда, постепенно углубляющаяся и разделяющая клетку на две половины - дочерние клетки, в каждой их которых имеется по ядру. У растений деление происходит путем образования так называемой клеточной пластинки, разделяющей цитоплазму: она возникает в экваториальной области веретена, а затем растет во все стороны, достигая клеточной стенки (т.е. растет изнутри кнаружи). Клеточная пластинка формируется из материала, поставляемого эндоплазматической сетью. Затем каждая из дочерних клеток образует на своей стороне клеточную мембрану и, наконец, на обеих сторонах пластинки образуются целлюлозные клеточные стенки. Особенности протекания митоза у животных и растений приведены в таблице 2.
Таблица 2. Особенности митоза у растений и у животных
| Растительная клетка | Животная клетка |
| Центриолей нет Звезды не образуются Образуется клеточная пластинка При цитокенезе борозда не образуется Митозы преимущественно происходят в меристемах | Центриоли имеются Звезды образуются Клеточная пластинка не образуется При цитокинезе образуется борозда Митозы происходят в различных тканях организма |
Так из одной клетки формируются две дочерние, в которых наследственная информация точно копирует информацию, содержавшуюся в материнской клетке. Начиная с первого митотического деления оплодотворенной яйцеклетки (зиготы) все дочерние клетки, образовавшиеся в результате митоза, содержат одинаковый набор хромосом и одни и те же гены. Следовательно, митоз - это способ деления клеток, заключающийся в точном распределении генетического материала между дочерними клетками. В результате митоза обе дочерние клетки получают диплоидный набор хромосом.
Весь процесс митоза занимает в большинстве случаев от 1 до 2 часов. Частота митоза в разных тканях и у разных видов различна. Например, в красном костном мозге человека, где каждую секунду образуется 10 млн эритроцитов, в каждую секунду должно происходить 10 млн. митозов. А в нервной ткани митозы крайне редки: так, в центральной нервной системе клетки в основном перестают делиться уже в первые месяцы после рождения; а в красном костном мозге, в эпителиальной выстилке пищеварительного тракта и в эпителии почечных канальцев они делятся до конца жизни.
Регуляция митоза, вопрос о пусковом механизме митоза.
Факторы, побуждающие клетку к митозу точно не известны. Но полагают, что большую роль играет фактор соотношения объемов ядра и цитоплазмы (ядерно-плазменное соотношение). По некоторым данным, отмирающие клетки продуцируют вещества, способные стимулировать деление клетки. Белковые факторы, отвечающие за переход в фазу М, первоначально были идентифицированы на основе экспериментов по слиянию клеток. Слияние клетки, находящейся в любой стадии клеточного цикла, с клеткой находящейся в М фазе, приводит к вхождению ядра первой клетки в М фазу. Это означает, что в клетке находящейся в М фазе существует цитоплазматический фактор способный активировать М фазу. Позднее этот фактор был вторично обнаружен в экспериментах по переносу цитоплазмы между ооцитами лягушки, находящимися на различных стадиях развития, и был назван "фактором созревания" MPF (maturation promoting factor). Дальнейшее изучение MPF показало, что этот белковый комплекс детерминирует все события М-фазы. На рисунке показано, что распад ядерной мембраны, конденсация хромосом, сборка веретена, цитокинез регулируются MPF.
Митоз тормозится высокой температурой, высокими дозами ионизирующей радиации, действием растительных ядов. Один из таких ядов называется колхицин. С его помощью можно остановить митоз на стадии метафазной пластинки, что позволяет подсчитать число хромосом и дать каждой из них индивидуальную характеристику, т. е. провести кариотипирование.
4. Амитоз. Эндорепродукция
Амитоз (от греч. а – отриц. частица и митоз) -прямоеделение интерфазного ядра путем перешнуровывания без преобразования хромосом. При амитозе не происходит равномерное расхождение хроматид к полюсам. И это деление не обеспечивает образование генетически равноценных ядер и клеток. По сравнению с митозом амитоз более кратковременный и экономичный процесс. Амитотическое деление может осуществляться несколькими способами. Наиболее распространенный тип амитоза – это перешнуровывание ядра на две части. Этот процесс начинается с разделения ядрышка. Перетяжка углубляется, и ядро разделяется надвое. После этого начинается разделение цитоплазмы, однако это происходит не всегда. Если амитоз ограничивается только делением ядра, то это приводит к образованию дву- и многоядерных клеток. При амитозе может также происходить почкование и фрагментация ядер.
Клетка, претерпевшая амитоз, в последующем не способна вступить в нормальный митотический цикл.
Амитоз встречается в клетках различных тканей растений и животных. У растений амитотическое деление довольно часто встречается в эндосперме, в специализирующихся клетках корешков и в клетках запасающих тканей. Амитоз также наблюдается в высокоспециализированных клетках с ослабленной жизнеспособностью или дегенерирующих, при различных патологических процессах, таких как злокачественный рост, воспаление и т. п.
Основным процессом в подготовке клетки к митозу является репликация ДНК и удвоение хромосом. Но синтез ДНК и митоз непосредственно не связаны, т.к. окончательный синтез ДНК не является непосредственной причиной вступления клетки в митоз. Поэтому в ряде случаев клетки после удвоения хромосом не делятся, ядро и все клетки увеличиваются в объеме, становятся полиплоидными. Такое явление - редупликация хромосом, без деления, выработалась в процессе эволюции как способ, обеспечивающий рост органов без увеличения числа клеток. Все случаи, когда происходит редупликация хромосом или репликация ДНК, но не наступает митоз, называются эндорепродукциями. Клетки становятся полиплоидными. Как постоянный процесс эндорепродукция наблюдается в клетках печени, эпителия мочевыводящих путей млекопитающих. В случае эндомитоза хромосомы после редупликации становятся видны, но ядерная оболочка не разрушается.
Если делящиеся клетки на некоторое время охладить или об работать их каким-либо веществом, разрушающим микротрубочки веретена (например, колхицином), то деление клеток прекратит ся. При этом исчезнет веретено, а хромосомы без расхождения к полюсам будут продолжать цикл своих превращений: они начнут набухать, одеваться ядерной оболочкой. Так возникают за счет объединения всех неразошедшихся наборов хромосом крупные новые ядра. Они, естественно, будут содержать вначале 4п число хроматид и соответственно 4с количество ДНК. По определению, это уже не диплоидная, а тетраплоидная клетка. Такие полипло идные клетки могут из стадии gi переходить в S -период и, если убрать колхицин, снова делиться митотическим путем, давая уже потомков с 4 п числом хромосом. В результате можно получить полиплоидные клеточные линии разной величины плоидности. Этот прием часто используется для получения полиплоидных растений.
Как оказалось, во многих органах и тканях нормальных ди плоидных организмов животных и растений встречаются клетки с крупными ядрами, количество ДНК в которых кратно больше 2 п. При делении таких клеток видно, что количество хромосом у них также кратно увеличено по сравнению с обычными дипло идными клетками. Эти клетки являются результатом соматиче ской полиплоидии. Часто это явление называют эндорепродук цией - - появление клеток с увеличенным содержанием ДНК. Появление подобных клеток происходит в результате отсутствия в целом или незавершенности отдельных этапов митоза. Суще ствует несколько точек в процессе митоза, блокада которых приведет к его остановке и к появлению полиплоидных клеток. Блок может наступить при переходе от С 2 -периода к собствен но митозу, остановка может произойти в профазе и метафазе, в последнем случае часто происходит нарушение целостности ве ретена деления. Наконец, нарушения цитотомии также могут пре кратить деление, что приведет к появлению двуядерных и поли плоидных клеток.
При естественной блокаде митоза в самом его начале, при переходе G 2 - профазы, клетки приступают к следующему циклу репликации, который приведет к прогрессивному увеличению ко личества ДНК в ядре. При этом не наблюдается никаких морфо логических особенностей таких ядер, кроме их больших размеров. При увеличении ядер в них не выявляются хромосомы митоти ческого типа. Часто такой тип эндорепродукции без митотической конден сации хромосом встречается у беспозвоночных животных, обнаруживается он также и у позвоночных животных, и у растений. У беспозвоночных в результате блока митоза степень поли плоидии может достигать огромных значений. Так, в гигантских нейронах моллюска тритонии, ядра которых достигают величины до 1 мм (!), содержится более 2-10 5 гаплоидных наборов ДНК. Другим примером гигантской полиплоидной клетки, образо вавшейся в результате редупликации ДНК без вступления кле ток в митоз, может служить клетка шелкоотделительной железы тутового шелкопряда. Ее ядро имеет причудливую ветвистую форму и может содержать огромные количества ДНК. Гигантские клетки железы пищевода аскариды могут содержать до 100000с ДНК.
Особый случай эндорепродукции представляет собой увеличе ние плоидности путем политении. При политении в S -периоде при репликации ДИК новые до черние хромосомы продолжают оставаться в деспирализованном состоянии, но располагаются друг около друга, не расходятся и не претерпевают митотическую конденсацию. В таком истинно интерфазном виде хромосомы снова вступают в следующий цикл репликации, снова удваиваются и не расходятся. По степенно в результате репликации и нерасхождения хромосомных нитей образуется многонитчатая, политенная структура хромосо мы интерфазного ядра. Последнее обстоятельство необходимо под черкнуть, так как такие гигантские политенные хромосомы ни когда не участвуют в митозе, более того - это истинно интерфаз ные хромосомы, участвующие в синтезе ДНК и РНК. От митотических хромосом они резко отличаются и по разме рам: в несколько раз толще митотических хромосом из-за того, что состоят из пучка множественных неразошедшихся хро матид - по объему политенные хромосомы дрозофилы в 1000 раз «больше митотических. Они в 70-250 раз длиннее митотических из-за того, что в интерфазном состоянии хромосомы менее конденсированы (спирализованы), чем митотические хромосомы. Кроме того, у двукрылых их общее число в клетках равно гаплоидному из-за того, что при политенизации происходит объединение, конъюгация гомологичных хромосом. Так, у дрозофилы в диплоидной соматической клетке 8 хромосом, а в гигантской клетке слюнной железы - 4. Встречаются гигантские полиплоидные ядра с политенными хромосомами у некоторых личинок двукрылых насекомых в клет ках слюнных желез, кишечника, мальпигиевых сосудов, жирового тела и т. д. Описаны политенные хромосомы в макронуклеусе инфузо рии стилонихии. Лучше всего этот тип эндорепродукции изучен у насекомых. Было подсчитано, что у дрозофилы в клетках слюнных желез может произойти до 6-8 циклов редупликации, что приведет к общей плоидности клетки, равной 1024. У некоторых хирономид (их личинку называют мотылем) плоидность в этих клетках до стигает 8000-32000. В клетках политенные хромосомы начинают быть видны после достижения политении в 64-128 п, до этого такие ядра ничем, кроме размера, не отличаются от окружающих диплоидных ядер.
Отличаются политенные хромосомы и своим строением: они структурно неоднородны по длине, состоят из дисков, междис ковых участков и пуфов. Рисунок расположения дисков строго характерен для каждой хромосомы и отличается даже у близких видов животных. Диски представляют собой участки конденсированного хро матина. Диски могут отличаться друг от друга по толщине. Общее их число у политенных хромосом хирономид достигает 1,5-2,5 тыс. У дрозофилы имеется около 5 тыс. дисков. Диски разделены междисковыми пространствами, состоящими, так же как и диски, из фибрилл хроматина, только более рыхла упакованных. На политенных хромосомах двукрылых часто видны вздутия, пуфы. Оказалось, что пуфы возникают на местах некоторых дис ков за счет их деконденсации и разрыхления. В пуфах выявля ется РНК, которая там же и синтезируется. Рисунок расположения и чередования дисков на политенных хромосомах постоянен и не зависит ни от органа, ни от возраста животного. Это является хорошей иллюстрацией одинаковости качества генетической информации в каждой клетке организма. Пуфы являются временными образованиями на хромосомах, и в процессе развития организма существует определенная последовательность в их появлении и исчезновении на гене тически различных участках хромосомы. Эта последо вательность различна для разных тканей. Сейчас доказано, что образование пуфов на политенных хромосомах - - это выражение генной активности: в пуфах синтезируются РНК, необходимые для проведения белковых синтезов на разных этапах развития насекомого. В естественных условиях у двукрылых особенно активны в отношении синтеза РНК два самых крупных пуфа, так называе мые кольца Бальбиани, который описал их 100 лет тому назад.
В других случаях эндорепродукции полиплоидные клетки воз никают в результате нарушений аппарата деления - веретена: при этом происходит митотическая конденсация хромосом. Такое явление носит название эндомитоз, потому что конденсация хро мосом и их изменения происходят внутри ядра, без исчезновения ядерной оболочки. Впервые явление эндомитоза было хорошо изучено в клетках: различных тканей водяного клопа - - геррии. В начале эндоми тоза хромосомы конденсируются, благодаря чему становятся хо рошо различимы внутри ядра, затем хроматиды обособляются, вытягиваются. Эти стадии по состоянию хромосом могут соответствовать профазе и метафазе обычного митоза. Затем хромосомы в таких ядрах исчезают, и ядро принимает вид обычного интер фазного ядра, но размер его увеличивается в соответствии с уве личением плоидности. После очередной редупликации ДНК такой цикл эндомитоза повторяется. В результате могут возникнуть полиплоидные (32 п) и даже гигантские ядра. Сходный тип эндомитоза описан при развитии макронуклеу сов у некоторых инфузорий, у целого ряда растений.
Результат эндорепродукции: полиплоидия и увеличение размеров клетки.
Значение эндорепродукции: не прерывается деятельность клетки. Так, например, деле ние нервных клеток привело бы к временному выключению их функций; эндорепродукция позволяет без перерыва в функциони ровании нарастить клеточную массу и тем самым увеличить объ ем работы, выполняемый одной клеткой.
увеличение продуктивности клеток.